Test test test
Archives
7. Cognitive adaptations
7.1. The cognitive niche hypothesis
Following our discussion on the general physical evolution of humans in the previous section, we now move on to an examination of how language are processed and understood. The cognitive niche hypothesis first advanced in Pinker & Bloom (1990) and later revised in Pinker (2010), is a consolidation of the human traits that distinguishes the natural language from other forms of animal communication. According to this framework, humans possess an innate mental state that provides them the basis for acquisition and knowledge of grammar, and to share it among themselves.
This cognitive niche includes the prerequisites of possessing a theory of mind, and also a need for social interaction. A number of scholars have proposed that the roots of theory of mind may be traced to mirror neurons and also the episodic memory in the primate brain, before it evolves into a distinctively human trait.
Though mirror neurons and episodic memory are found to be of separate systems in the human brain, both are able to give us a broader contexts for understanding the complementary functions of the human language and mind.
[Back to Table of Contents]
7.2 Mirror neurons
Mirror neurons are a class of neurons which are activated as an animal executes a movement via motor control and when they imitate another individual’s (possibly conspecific) behaviour (Kilneremail & Lemon, 2013). These mirror neurons are hypothesized to be an adaptation mechanism for the understanding of actions, imitation-learning, and of associative learning. Perception and the genetic basis of human vocal imitation are suggested to be linked by mirror neurons similar to that of the monkeys in a way that is connected to the reciprocity of linguistic signs. This hypothesis is supported by some homologies discovered between the Macaque monkey’s premotor area F5 and also the human Broca’s area, as seen in Figure 5 below.

Figure 7: Mirror Neurons in the premotor cortex of macaque monkeys (indicated by red and yellow areas in A above) and in the right superior temporal sulcus of humans (indicated by red and yellow areas in B above)
Studies with Magnetic Resonance Imaging (MRI) have discovered that some cortical areas of the human brain like the inferior frontal cortex (near to the Broca’s area) are homologous to the monkey mirror neuron system. As the Broca’s area is thought to be one of the most language centered regions of the brain, one possible hypothesis is that the human language may have been evolved from the system of mirror neurons. The distinction of the mirror neuron system between monkeys and humans is that the human mirror neurons are able to be activated by linguistically related actions like reading and listening.
Human children have the ability to vocally repeat pseudowords via echolalia and speech shadowing. Such speech repetition and imitation not only helps the children to acquire new sets of vocabulary, this process of language acquisition via speech repetition is instinctive and can occur without comprehension and understanding. This suggests that the development of mirror neurons is a feature associated with a strong genetic predisposition of humans, favored by natural selection.
It is plausible that genetically pre-programmed mirror neurons was a result of a gradual process of language evolution. Improvements in cognitive mechanisms could have been selected for the mirror neurons’ ability to recognize, imitate and familiarise with complex actions. This function may account for the influence of prefrontal cortex and ventral pathways. Though there may not be concrete evidence that mirror neurons are a genetic adaptation specifically humans’, it remains that genetics may dictate the learning process of neurons and not just what they learn.
Alternatively, there is also the theory that mirror neurons are a result of associative learning instead of evolutionary adaptation. It suggests that the properties of the mirror neurons are a product, not of a genetic predisposition, but of the general processes of associative learning like conditioning procedures. The associative hypothesis conjectures that mirror neurons can also be found in other non human species. For instance, the quality of speech sounds known as formants are perceivable by many vertebrates such as primates and birds. The ability to accurately distinguish formants specific to different individuals of their species suggests that mirror neurons may also be present in animals. Specifically, a related example is the honeybees. The honeybees are able to make use of associative learning to distinguish individual human faces. When the honeybees are placed in an environment whereby the recognition of human faces are crucial, associative learning are optimized for the foraging behavior of the honeybees. Given that human faces were not originally part of the environment where the honeybee nervous systems evolved, associative learning is not an adaptation for face discrimination in honeybees (Cook et. al, 2014).
However, the associative learning is not sufficient to account for the development of mirror neurons. For instance, genetic predispositions interacts with associative learning; the associative account is unable to justify the reason why some behaviors are learnt more easily than others. Genetic predispositions hence may interact with associative learning to produce efficiency in language learning. Many experiments have also shown that human infants are inclined to imitate humans as opposed to non human actions (Bertenthal, 2014). Thus, associative learning is necessary but not an sufficient enough explanation for mirror neuron development.
It remains that the discovery of mirror neurons have result in breakthroughs in the gestural theory of speech origin. Mirror neurons in humans facilitates a direct communication between interlocutors. The actions communicated by an individual can be understood by an observer as it is able to elicit the same motor representation in their parieto-frontal mirror system. It is proposed that the mirror neurons are the basic mechanism where human language evolved. This is because it can be used to explain one of the fundamental difficulties for understanding language evolution- how messages conveyed can become valid for both the interlocutors.
7.3 Episodic memory
According to Tulving (1972), episodic memory is an important cognitive development unique to humans. Controlled by the prefrontal medial temporal regions and posterior regions such as the posterior cingulate and retrosplenial cortex, it refers to the ability of humans to remember and re-experience past events (episodic memory) and to anticipate future events. Concerned with conscious recollection of specific past experiences, episodic memory allows humans to ‘travel’ mentally in time is important for encoding the ability for learning and retaining language skills.
This ability may have co-evolved as an adaptation with the genus Homo millions of years ago, as it allows the documentary of important historical and environmental events (cave paintings etc) , giving humans the ability to analyse and compare present, past and future scenarios.
Some features of the grammatical language may have also been derived from this pressure to communicate the timing of events (displacement) so as to strategize and enhance group survival in difficult terrains and environments. Thus this communicative cooperation amongst humans with regards to future references and planning of events may have co-evolved with symbolic communication systems.
Episodic memories are also generative, thus resulting in the productivity and creative attribute of human language. When strategizing for future events, humans have the capacity to remember and compare the different scenarios from their store of semantic concepts and episodic memories (Corballis, 2010).
Another point to note is that, more recent research has shown that there are at least some animals, notably a few primates and corvids, that exhibits the potential for episodic memory, and thus it may not be an aspect that is unique only to humans. However, more extensive research is needed to support this theory.

6. Physical adaptations
Now that we know the general idea of the theory of the evolution of humans, let us discuss some of the physical prerequisites for language in humans.
6.1 Descent of the larynx
Most languages are spoken in their most basic form and thus humans require the ability to pronounce consonants and vowels. This ability is only found in humans and n
7.1.1 The Human descended larynx
From William T. Fitch’s speech on the Evolution of Language (2005), the larynx, also known as the “voice box” for humans, can engage into the nasal cavity of animals, allowing the ease of breathing while swallowing simultaneously.The larynx in humans is lower than that of other primates, allowing more space for the the tongue to move during speech production.The descended tongue root (i.e. the back of the tongue) allows additional degree of freedom for vocal tract acrobatics during speech production. This mobility of the tongue allows it to produce highly coordinated motions, and therefore giving humans the ability to produce consonants and vowels. The difference in anatomy of other primates’ vocal tracts from humans thus prevent them from speaking well, even with speech training.
6.1.2 The comparative method on descent of the larynx
A descended larynx was believed to be uniquely human until recently. However, it has been shown that animals generally lower their larynx during vocalization (e.g. dog barking) when vocalizing (i.e. producing sounds). Lowering of the larynx during vocalization may hence, be an analogous trait (i.e. traits that are similar in function, but from a different evolutionary source).
Additionally, several species of animals have similarly been observed to possess descended larynx. Examples include koalas, deers and even lions. Since the common mammalian ancestor of these species do not have a descended larynx, this convergence indicates that selective evolution may have taken place. From this discovery, Fitch (2000) proposed that the motivation for the descent of the larynx was not only caused by the need for speech in humans, since this descended larynx in animals did not cause them to speak, unlike the descended larynx in humans.The evolution of the vocal tract thus requires consideration of other factors like selective evolution. Both homology (i.e. similar function, but different evolutionary source) and analogy traits must then be analysed for a comprehensive understanding of human’s capacity for speech.This comparative approach presented by Fitch (2005) then suggests a probable hypothesis that the trait of a lowered larynx may have already been present even before the the humans and chimpanzees species split from a possible common ancestor.Further research was then done on nervous and muscular control to explore other prerequisites for language to account for the reason behind this occurrence. Specific movements of mouth and teeth are crucial to the production of consonants and vowels. Thus, researchers hypothesised that there could be a particular gene in humans that control this fine motor skill, leading to the significance of the research on the FOXP2 gene.
[Back to Table of Contents]
6.2 Nervous and muscular control – FOXP2
Another physical prerequisite for language in human beings is the presence of the FOXP2 gene. In general, FOXP2 in humans is a gene which codes for oral motor control and speech production in humans. Speech production in human beings require fine nervous control. One of the most evident aspect is that enhanced motor control over vocal articulators like the tongue, lips, jaw, etc., is needed for speech production. For example, the movements of the tongue has to be synchronised with the lips and vibrations of the larynx (voice box) when we speak (Liberman & McKarthy, 2007).
It is important to note that the above discussion on FOXP2 applies to the human variety of the gene. However, since this blog focuses on the features and origins of human language, we will not discuss how this gene codes for other functions in vertebrates other than humans. More information on the differing function of the FOXP2 gene in animals can be found in this this hyperlink.
To conclude this section, both the descent of the larynx, and the presence of FOXP2 in humans are crucial for speech production, and thus are also important in the physical development of humans to produce language. However, in terms of processing and understand
[Back to Table of Contents]
5. Phylogenesis of humans
5.1 The co-evolution of humans and language
Much research on the evolution of language has asserted that the phylogenesis, or evolutionary descent, of the homo sapiens species has played a key role in the emergence of language, and not simply because modern humans are language’s sole users (Chater and Christiansen, 2011; Kirby, Cornish & Smith, 2008; Pinker, 2003).
The following figure 3 from Christiansen and Kirby (2003) provides a concise snapshot of the co-evolution of humans and language, as an interaction between three different evolutionary systems: biological evolution (phylogeny), individual differences in learning (ontogeny) and rate of language change.

Figure 3: the co-evolution of humans and language (Christiansen & Kirby, 2003)
Accordingly, by charting the physical and cognitive evolutionary developments in humans, we expect to glean crucial insights into the phylogenesis of language itself. Let’s first begin with a survey of the general phylogenetic history and evolutionary stages of homo sapiens.
[Back to Table of Contents]
5.2 Phylogenetic branching leading to homo sapiens
According to Fitch (2007), the ‘independent evolutionary history’ of humans starts from the last common ancestor (LCA) between chimpanzees and hominids, around ‘seven million years ago’. Hominids, are a sub-branch of primates, and include all known human species, although only one hominid species exists today (i.e. all modern humans), namely homo sapiens.
The following figure pictorially charts the phylogenesis of humans as a specific species of hominids.
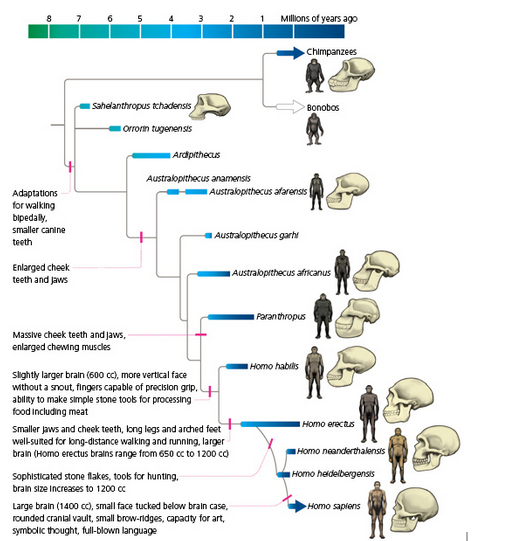
Figure 5: The Phylogeny of Humans as adapted from Evolution 101
Additionally, within the hominid branch, two large sub-groups can be located; namely, the earlier genus Australopithecus (includes notably the specific species of anamensis, afarensis, africanus, boisei, robustus) and the later genus Homo (includes notably the specific species of habilis, ergaster, erectus, neanderthalensis, sapiens).
[Back to Table of Contents]
5.3 Adaptive stages leading to homo sapiens
5.3.1 The fossil data record – three stages of hominid adaptations
Given this general phylogenetic picture, palaeontologists have attempted to reconstruct in greater detail the specific evolutionary pathway of modern humans from fossil evidences. Accordingly, some researchers have suggested that the language-using Homo sapiens species emerged from three broad stages of hominid adaptation (Fitch, 2007), which are detailed below.
5.3.2 Adaptation stage 1: Bipedalism
The first major hominid evolutionary adaptation occurred roughly six million years ago (6 mya), in the early human species Australopithecus farensis. Palaeontological analysis on a fossil of this pre-homo hominid species named ‘Lucy’ found clear skeletal evidence for an upright posture. This, in turn, indicated the presence of bipedalism – defined as the mode of locomotion whereby an organism moves by means of its two rear limbs or legs – in this species.
Many scholars believe that the evolution of bipedalism had a huge impact on the cognitive development of later human species (after Australopithecus afarensis). By reducing the need for four-limbed locomotion (e.g. climbing), later human species would have faced a significantly different visual representation of cognitive space and distance, compared to earlier non-bipedal hominids. Further, some scholars also draw a link between the role of imitation systems (mirror neurons, to be elaborated in section 6: cognitive prerequisites) as a primary selective pressure for bipedal locomotion, thus linking bipedalism, imitative abilities and language-learning abilities in an evolutionary chain. Practically and finally, an upright posture would have freed up hands of later hominids to engage in other activities – such as in communicative gestures or tool-making – which has been conjectured to have led to the second major stage of hominid adaptation: brain expansion.
5.3.3 Adaptation stage 2: Basic tool making and Brain expansion
The second major hominid evolutionary adaptation is estimated to have started around the time of the genus Homo. This involved the expansion of the human brain – the neocortex – which is, on average, around 35-60% larger in proportion to a typical primate’s body and brain size. Various scholars have suggested a period just under 4 million years, a relatively short time in evolutionary terms, in which the hominid brain had grown about three times its original size.
Many hypotheses have been made regarding the developmental effects of this brain expansion. This have included suggestions that such a physical adaptation could have resulted in various behavioural peculiarities in humans, such as a theory of mind, intelligence, and other cognitive prerequisites that may account for the human language, including the development of more acute motor skills and greater cognitive capacity.
In other words, this expansion of brain was probably crucial for the development of language. As earlier noted, this adaptation was had occurred quite early – at or near the beginning of the genus Homo itself – specifically in the Broca’s area, the part of the brain closely associated with language. Fossil evidences have helped paleontologists trace the presence of Broca’s area in endocasts from various genus Australopithecus. Wernicke’s area was discovered in fossil records later on, amongst the Homo habilis (Dubuc, n.d).
5.3.4 Adaptation stage 3: Advanced tool making and Motor control
The rise of tool-making, following the freeing of hands due to bipedalism, triggered off the third major stage of hominid evolutionary adaptation.
The development of sophisticated tools is considered a crucial moment in the human evolution, as it set the stage for better nutrition and advanced social behaviours, like the planning and division of labour and group hunting, reinforcing the notion that tool making and language evolved together as both required more complex and advanced cognition.
The parallel of the complexity in the tools made by humans and development of language may imply a correlation between enhanced motor movement and an early form of the human language used to communicate thoughts and intentions. For instance, the design of stone tools have evolved and advanced quickly in the human pre-history, beginning with sharp and primitive stone flakes, and culminating in finely honed and produced hand axes.
Notably, deficits in motor control are also often linked to aphasia in humans. Since there is a strong correlation between manual tool making and speech communication, the investigation on the evolution of humans’ physical and cognitive characteristics and their specific changes offers great promise on ground discussions of language evolution.
[Back to Table of Contents]
References
Bertenthal, B. I. (2014). The insufficiency of associative learning for explaining development: Three challenges to the associative account. BEHAVIORAL AND BRAIN SCIENCES (2014). doi:10.1017/S0140525X1300221
Connor, R. C. (2007, 04). Dolphin social intelligence: Complex alliance relationships in bottlenose dolphins and a consideration of selective environments for extreme brain size evolution in mammals. Philosophical Transactions of the Royal Society B: Biological Sciences, 362(1480), 587-602. doi:10.1098/rstb.2006.1997
Cook, R., Bird, G., Catmur, C., Press, C., & Heyes, C. (2014). Mirror neurons: From origin to function. BEHAVIORAL AND BRAIN SCIENCES, 177–241. doi:10.1017/S0140525X13000903
Corballis, M. C. (2010). Mirror neurons and the evolution of language. Brain And Language, 112(1), 25-35. doi:10.1016/j.bandl.2009.02.002
Dubuc, B. (n.d.). The Brain from Top to Bottom. Retrieved November 2014, from http://thebrain.mcgill.ca/intermediaire.php
Fishlock, V., Lee, P. C. (2013). Forest Elephants: Fission-Fusion and Social Arenas. Animal Behaviour, 85, 357-363. Retrieved from: https://www.sciencedirect.com/science/article/pii/S0003347212005143?via%3Dihub
Fitch, W. T. (2000). The evolution of speech: a comparative review, Trends in Cognitive Sciences 4(7): 258-267.
Fitch, W. T. (2005). The evolution of language: a comparative review. Biology and philosophy, 20(2-3), 193-203.
Fitch, W. T. (2007). The Evolution of Language: A Comparative Perspective, in Oxford Handbook of Psycholinguistics, edited by G. Gaskell. Oxford University Press, Oxford.
Givón, T. (2002) Bio-Linguistics: The Santa Barbara Lectures. Benjamins: Amsterdam.
Hedeager, U. (n.d.). IS LANGUAGE UNIQUE TO THE HUMAN SPECIES? Retrieved from http://www.columbia.edu/~rmk7/HC/HC_Readings/AnimalComm.pdf
Hockett, C. F. (1960). The Origin of Speech, Scientific American 203: 88–111. Reprinted in: Wang, William S-Y. (1982) Human Communication: Language and Its Psychobiological Bases, Scientific American, pp. 4–12
Jenkins, L. (1999). Biolinguistics: Exploring the Biology of Language. Cambridge University Press, New York.
Kershenbaum, A. E. Bowles, T. M. Freeberg, D. Z. Jin, A. R. Lameira, K. Bohn. Animal vocal sequences: not the Markov chains we thought they were. Proceedings of the Royal Society B: Biological Sciences, 2014; 281 (1792): 20141370 DOI: 10.1098/rspb.2014.1370
Kilneremail, J. M., & Lemon, R. N. (2013). What We Know Currently about Mirror Neurons (MiniReview). Retrieved from http://dx.doi.org/10.1016/j.cub.2013.10.051
Kirby, S. (2002). Natural Language From Artificial Life. Retrieved from http://www.lel.ed.ac.uk/~simon/Papers/Kirby/Natural%20Language%20from%20Artificial%20Life.pdf
Kirby, S., Cornish, H., & Smith, K. (2008). Cumulative cultural evolution in the laboratory: An experimental approach to the origins of structure in human language. Proceedings of the National Academy of Sciences of the United States of America 105 (31): 10681-10686
Kuthy, K. D. (2001, September 28). Arbitrariness in Language. Retrieved from http://www.ling.ohio-state.edu/~kdk/201/autumn01/slides/arbitrariness-4up.pdf
[VID] William T. Fitch: The Evolution of Language http://www.youtube.com/watch?v=eP0Kdkz4iHU
Pinker, S. (2010). The cognitive niche: Coevolution of intelligence, sociality, and language. Proceedings of the National Academy of Sciences 107 (2): 8993–8999
Understanding Evolution 101. (n.d.). Retrieved from http://evolution.berkeley.edu/evolibrary/home.php
Part II: The Evolution of Language

How did language emerge?
4. Animal Language
4.1 Animal Language
While the name “Animal Language” might be a little of misnomer since there has not been a single animal language that manages to fulfill all of Hockett’s design feature (even excluding the ones that were deemed as unnecessary in definition above), it serves as a good collective name for animal communication as a whole, and allows for easy comparison with what we term human language. However, it is important to note that animals can convey various message to each other. For example, to give warnings or to express their emotions such as anger and fear. In addition, animals communicate to locate food sources as well as their desire to willingness to mate. Thus, a more accurate term for animal language would be animal communication. The communicative capacity of animals is still not able to match the sophistication and productivity of human language. This section thus explores the communicative ability of animals, contrasting it with features of human language, as designated by Hockett.
[Back to Table of Contents]
4.2 Comparison with human language
Since the 1960s when Hockett first described a set of 13 features defining human language and how these features putatively made it distinct from other forms of animal communication, advances the study of animal communication have also revealed closer similarities with some of Hockett’s design features of language.
We’ve seen that animals do have their own unique communication systems, the closest to human language being animal calls. There are several types of calls. There are contact calls and food calls to help coordinate group movements, as seen with dolphins, bats, and pinnipeds, which have individually-specific calls or whistles. There are also softer calls used in familial or intimate contexts, as seen with altricial birds, primates, and marmoset baby babbling.
The evolution of vocal imitation in humans, some birds, elephants, bats, and dolphins may be a result of needing to use acoustic signals to maintain individual-specific bonds when animals separate and reunite in fission-fusion societies.
4.2.1 Honeybees
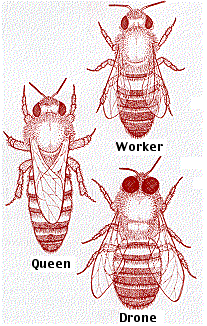 Apis mellifera commonly known as the domestic honey bee is a colonial insect living in hives that are made up of one queen (a fertile female), a few drones (males) and thousands of workers (infertile females). The workers are responsible for foraging for food namely nectar and pollen as well as tending to the young (2014). German ethologist Karl von Frisch’s (1886–1982) theorized that bees communicate the distance and the direction of food sources to each other via two kinds of dances: the round dance and tail-waggle dance (Munz, T. 2005), therefore indicating that bees exhibit a form of displacement when communicating with each other.
Apis mellifera commonly known as the domestic honey bee is a colonial insect living in hives that are made up of one queen (a fertile female), a few drones (males) and thousands of workers (infertile females). The workers are responsible for foraging for food namely nectar and pollen as well as tending to the young (2014). German ethologist Karl von Frisch’s (1886–1982) theorized that bees communicate the distance and the direction of food sources to each other via two kinds of dances: the round dance and tail-waggle dance (Munz, T. 2005), therefore indicating that bees exhibit a form of displacement when communicating with each other.
4.2.1.1 Round Dance
 Round dance is usually performed when bees find food source which are less than 50 meters from their hives. Bees will run around in narrow circles before reversing direction to their original course. This may be repeated a few times at the same location or move to another location on the comb. Although a round dance communicates distance, it does not reflect the direction (R. Tarpy, D., n.d.).
Round dance is usually performed when bees find food source which are less than 50 meters from their hives. Bees will run around in narrow circles before reversing direction to their original course. This may be repeated a few times at the same location or move to another location on the comb. Although a round dance communicates distance, it does not reflect the direction (R. Tarpy, D., n.d.).
4.2.1.2 Waggle Dance
 Bees will perform the waggle dance in the shape of figure-eights when returning from sources which are located farther than 75 meters from their hives. Von Frisch mentioned that the frequency of turns in the dances varied inversely with the distance of foods while the straight runs of the dances indicated their direction of the food. He also found out that bees can detect polarized light and thus, oriented their dances and foraging flights with respect to the sun (Munz, T., 2005). Due to the directional information that is based upon the sun’s position, a forager’s dance for a particular resource will change during a day.
Bees will perform the waggle dance in the shape of figure-eights when returning from sources which are located farther than 75 meters from their hives. Von Frisch mentioned that the frequency of turns in the dances varied inversely with the distance of foods while the straight runs of the dances indicated their direction of the food. He also found out that bees can detect polarized light and thus, oriented their dances and foraging flights with respect to the sun (Munz, T., 2005). Due to the directional information that is based upon the sun’s position, a forager’s dance for a particular resource will change during a day.
4.2.2 Vervet Monkeys
 In addition, warning calls by the vervet monkey exhibit some level of semanticity as they produce distinctly different calls depending on the type of predator seen (Vagell, 2011a; 2011b). According to a 1980 experiment by Robert Seyfarth, Dorothy Cheney and Peter Marler, alarm calls for different types of predators sounded distinctly different and each call was able to elicit a distinct defensive response. When the leopard call was played, the subjects ran up into the trees while when eagle call were played, subjects looked up and run out of trees into lower bushes. When snake calls were played, subjects immediately stood on their hind legs and looked on the ground around them. Their calls however, show a lack of productivity as they only make warning calls that reflect the present situation and not in the past.
In addition, warning calls by the vervet monkey exhibit some level of semanticity as they produce distinctly different calls depending on the type of predator seen (Vagell, 2011a; 2011b). According to a 1980 experiment by Robert Seyfarth, Dorothy Cheney and Peter Marler, alarm calls for different types of predators sounded distinctly different and each call was able to elicit a distinct defensive response. When the leopard call was played, the subjects ran up into the trees while when eagle call were played, subjects looked up and run out of trees into lower bushes. When snake calls were played, subjects immediately stood on their hind legs and looked on the ground around them. Their calls however, show a lack of productivity as they only make warning calls that reflect the present situation and not in the past.
Here is a video that summarizes Seyfarth, Cheney and Marler’s (1980) research:
https://www.youtube.com/watch?t=188&v=3lsF83rHKFc
4.2.3 Elephants
 In 2006, keepers at Seoul’s Everland zoo were surprised when they heard Kosik, a 16-year-old elephant, talk. He had learned to talk by putting its trunk in its mouth and mimicking the words it heard from its caretakers: “yes,” “no,” “lie down,” “sit down,” and four other phrases in Korean. Scientists say that through Kosik they might be able to prove that elephants are capable of learning language, although it is unlikely that he understands the meaning of the words he expresses (Ilbonito, 2015). Such an occurrence is considerably rare and usually elephants will communicate by using gestures as well as “infrasounds” as explained below.
In 2006, keepers at Seoul’s Everland zoo were surprised when they heard Kosik, a 16-year-old elephant, talk. He had learned to talk by putting its trunk in its mouth and mimicking the words it heard from its caretakers: “yes,” “no,” “lie down,” “sit down,” and four other phrases in Korean. Scientists say that through Kosik they might be able to prove that elephants are capable of learning language, although it is unlikely that he understands the meaning of the words he expresses (Ilbonito, 2015). Such an occurrence is considerably rare and usually elephants will communicate by using gestures as well as “infrasounds” as explained below.
4.2.3.1 Gestures
 Biologist and conservationist Joyce Poole and her husband, Petter Granli, both of whom direct ElephantVoices, a charity they founded to research and advocate for conservation of elephants in various sanctuaries in Africa, have developed an online database decoding hundreds of distinct elephant signals and gestures. Poole and Granli have also deciphered the meaning of acoustic communication in elephants. Below are two forms of gestures that elephants commonly perform.
Biologist and conservationist Joyce Poole and her husband, Petter Granli, both of whom direct ElephantVoices, a charity they founded to research and advocate for conservation of elephants in various sanctuaries in Africa, have developed an online database decoding hundreds of distinct elephant signals and gestures. Poole and Granli have also deciphered the meaning of acoustic communication in elephants. Below are two forms of gestures that elephants commonly perform.
 Aggression: Normally in such an aggressive stance, an elephant will hold its head well above its shoulders and with tusks lifted, direct its gaze at its provoker. An elephant may also increase its height by standing on a log or an anthill to assume greater stature, an approach used by males when they’re sizing each other up.
Aggression: Normally in such an aggressive stance, an elephant will hold its head well above its shoulders and with tusks lifted, direct its gaze at its provoker. An elephant may also increase its height by standing on a log or an anthill to assume greater stature, an approach used by males when they’re sizing each other up.

- Death: Elephants are empathetic towards one another by holding funerals for the dead (Wolchover, 2012). Elephants will try to rouse an injured or fallen elephants by using their tusks and trunk to try and feed a dead elephant, or appear to lift or even carry sick, dying, or dead elephants (Poole et al., 2015).
4.2.3.2 Infrasounds
 A team of voice researchers and biologists led by Christian Herbst, Angela Stoeger and Tecumseh Fitch has discovered that elephants produce “infrasounds” which are very low frequency sounds, far from the hearing compatibility of the average human being. One possibility as to how “infrasounds” are produced is when the elephants tense and relax the muscles in their larynx for each pulse of sound. Another possibility is that they are produced like human speech or singing. Because the elephant larynx is so large, they are extremely low in frequency. The “infrasounds” can travel several kilometers and provide elephants with a “private” communication medium that plays a pivotal role in elephants’ complex social life (2012).
A team of voice researchers and biologists led by Christian Herbst, Angela Stoeger and Tecumseh Fitch has discovered that elephants produce “infrasounds” which are very low frequency sounds, far from the hearing compatibility of the average human being. One possibility as to how “infrasounds” are produced is when the elephants tense and relax the muscles in their larynx for each pulse of sound. Another possibility is that they are produced like human speech or singing. Because the elephant larynx is so large, they are extremely low in frequency. The “infrasounds” can travel several kilometers and provide elephants with a “private” communication medium that plays a pivotal role in elephants’ complex social life (2012).
4.2.3.3 Elephant Calls in Fission-Fusion Societies
African Savannah elephants belong to a complex fission-fusion society, and vocal imitation is a useful form of acoustic communication which helps maintain individual-specific bonds without changing social groupings. Two studies conducted on these elephants lead to interesting instances of vocal imitation. The first case involved a ten-year-old adolescent female African elephant living in a semi-captive group of orphaned elephants in Kenya. Trucks could sometimes be heard from the elephant’s enclosure which was 3 kilometres away from the highway, and it was found that she emitted truck-like sounds that were different from the normal calls of African elephants. The second case involved a 23-year-old male African elephant who spent 18 years with 2 female Asian elephants in a zoo in Switzerland, and he was found to be producing chirping sounds typically produced by Asian elephants but not African elephants. It strengthens the idea that communicative demands of maintaining social relationships in fluid societies can lead to vocal learning (Fishlock & Lee, 2013).
4.2.4 Dolphins
Dolphins are highly sociable creatures, with a large brain relative to their body mass. Looking at the figure below, we see how the expansion of the dolphin’s brain almost mirrors that of human beings – with the introduction of a form of communication and socializing propelling it to growth.
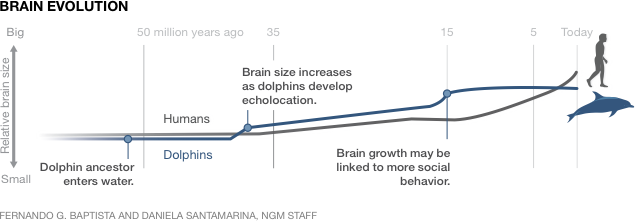
Most dolphins produce a variety of sounds described as clicks, whistles, buzzes, squawks, screams and barks (Herzing, 1996). Half a century of research has been dedicated to trying to decipher the languages that dolphins use, with not much progress. Scientists have not been able to determine the units that make up what might be an actual dolphin language. In recent times however, technology has allowed for the advancement of such research. Scientists have invented CHAT (cetacean hearing and telemetry), a machinery designed to not only record dolphins, but also produce their signature sounds as well other sounds that are dolphin-liked in nature. While at present, we are still in the dark about the possible language that the dolphins might hold, we know a decent amount of what their communication is capable of.
4.2.4.1 Clicking Noises In Isolation
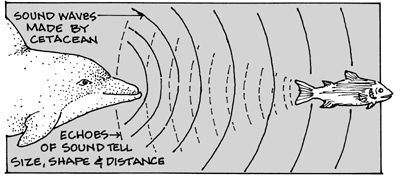 The echolocation sounds are short, broadband pulses (Evans, 1973) which are produced in low repetition rate trains or in high repetition rate bursts. Bursts of pulses can be used during prey capture (Verfuss et al., 1996) and during hostile interactions (Blomqvist & Amundin, 1998). The bottlenose dolphin uses echolocation clicks with a peak frequency between 115 and 121 kHz when recorded in open waters (Au, 1993). This mode of communication allows dolphins to navigate, recognize their friends and foes as well as escape from obstacles (Mary Cerullo, 2013).
The echolocation sounds are short, broadband pulses (Evans, 1973) which are produced in low repetition rate trains or in high repetition rate bursts. Bursts of pulses can be used during prey capture (Verfuss et al., 1996) and during hostile interactions (Blomqvist & Amundin, 1998). The bottlenose dolphin uses echolocation clicks with a peak frequency between 115 and 121 kHz when recorded in open waters (Au, 1993). This mode of communication allows dolphins to navigate, recognize their friends and foes as well as escape from obstacles (Mary Cerullo, 2013).
4.2.4.2 Whistling
 Whistles consist of narrow-band tones of constant frequency (CF) or tones that vary in frequency. Harmonic components are normally present (Popper, 1988). Dolphins use whistles to maintain contact within their pod or when meeting other pods of dolphins. Their whistles may indicate danger or a cry for help. Scientists think that each dolphin has its own unique whistle, almost akin to our names. Whistles may also help dolphins hunt cooperatively and coordinate migratory movements.
Whistles consist of narrow-band tones of constant frequency (CF) or tones that vary in frequency. Harmonic components are normally present (Popper, 1988). Dolphins use whistles to maintain contact within their pod or when meeting other pods of dolphins. Their whistles may indicate danger or a cry for help. Scientists think that each dolphin has its own unique whistle, almost akin to our names. Whistles may also help dolphins hunt cooperatively and coordinate migratory movements.
The whistles are used as a contact call when animals are out of visual range, and the possibilities of dolphin communication is made greater when combined with their imitation ability. Reports have been made that in Sarasota Bay, Florida, 43% of male calves and 10% of female calves developed signature whistles that were very similar to their mothers, but the question of whether signature whistle imitation in bottlenose dolphins is context-dependent remains inconclusive.
Herman in 2002 and 2006 trained bottlenose dolphins, a fission-fusion species to perform novel synchronous behaviours on command, and the study showed high levels of imitation with these dolphins. Interestingly, considerable research effort has failed to produce comparable skills in most non-human primates (Connor, 2007).
4.3 Further Research
Even though much research has been done on animal communication, more can be done to understand how and why different modes of vocalizations come about. One way to do so is to study the behaviour of animals in tightly knit social groups, such as dolphins and whales.
According to biologist Kathy Heise, resident killer whale communication is distinctly different from transient killer whales. The mode of communication employed by one group of whales does not seem to be mutually intelligible with the other. Future research on these “dialects” differing from family to family could help shed light on why dialect groups exist in human languages.
https://www.youtube.com/watch?v=-7E7FTSsbkg
To date, humans have not been able to fully bridge the gap with naturally sociable animals such as whales. Instead of trying to teach animals human language to understand how animals process language, humans could try to imitate and manipulate the sounds of animal vocalizations to see how these animals react to it. By manipulating and observing how animals react to these stimuli could help us eventually understand the exact meaning of their vocalizations. During a joint project by Australian telecommunications company Optus and the Humpback Acoustic Research Collaboration, a chamber orchestra was placed out in the sea while imitating sounds of a whale song, drawing the attention of curious humpback whales (Macleod, 2009). This shows us that there is the possibility of employing music as a communicative tool to bridge the language gap between animals and humans in order to shed some light on the acoustic properties of language.
[Back to Table of Contents]
4.4 Conclusion
Although there are some animals which seem to exhibit some forms of human language, animal and human communication are still a major contrast from each other. In the case of Kosik, for example, the question of whether the elephants are capable of expressive speech (as opposed to straight mimicry) has yet to be fully determined. .In conclusion, the use of human language is only unique to us and not only does it allow us to communicate with one another, it also allows us to form sentences with infinite variety and nonsensical yet understandable sentences.
A monkey can scream to warn others of an approaching predator, or alert them to tasty food source, but it cannot communicate something like “Doesn’t that hawk have a funny looking beak?” or “With a little salt, this fig would taste divine”. Jason G. Goldman, a science writer based in Los Angeles puts it simply:
Only humans are able to utter grammatical nonsense
While we continue to do further research as well as encounter new discoveries about animal communication, we seem to further and further establish the idea that our language – human language – is unique and much more expressive than any other form of communication found in the animal kingdom. We continue to attempt to teach animals our language, in a hope that one species will be able to adapt to it as well as we have, but continuously fall short in that regard. All signs seem to point towards the uniqueness of our language. However, as technology continues to improve, we begin to have more and more ways to decipher the current communication systems they have instead of teaching them our language. And maybe, just maybe, one of them will prove to us that we are not the only living creatures capable of what we consider to be language.
Of course, human language did not spring up suddenly, fully formed and fully versatile. The capacity for human language comes from a long line of evolution, with various stages and adaptations that all work together to form what we know today as Human Language. In the next part of the chapter, we will look at the evolution of human language; specifically, the evolution of the capacity of human language we now possess.
[Back to Table of Contents]
3. A Comparative Approach
3.1 Re-cap of existing studies on the nature of language
In section 1, we identified six core features of language which appeared to distinguish it as a uniquely human system of communication: Arbitrariness, Discreteness, Displacement, Productivity, Cultural Transmission and Duality of Patterning. In section 2, we identified three main components of language: a store of signals, rules for structuring these signals, and communally-agreed upon interpretations of these signals.
In this section, we put forward our own analysis of human language, which attempts to relate these two existing perspectives, in order explain how the structure of language components gives rise to such perceived features.
3.2 System: Two basic rule sets – Duality of Patterning
Firstly, every spoken language is made up of a set of speech sounds called phonemes. These phonemes, follow a set of systematic and language-specific rules, combine to form meaningful units called morphemes. These morphemes, in turn, follow another set of systematic and language-specific rules for combining into individuated words and sentences.
In other words, we can distinguish between two sets systematic rules at an abstract level, which in terms of Fitch’s componential view would be a distinction between the rules for structuring individually meaningless language signals directly versus the rules for structuring meaningful groups of language signals. Consequently, this helps explain the core feature of duality of patterning in natural languages.
3.2.1 Linguistic signs – Arbitrariness
Secondly, it is crucial to note that the first set of structuring rules which combine speech sounds into meaningful units are not semantically motivated. This is to say there is no logical relationship between, for example, English’s combination of the phonemes [d], [ɔ] and [g], to form the morpheme dog, and the idea of the specific animal it represents. Consequently, this gives rise to the core language feature of arbitrariness.
Arbitrariness is crucial in the human language because it frees up the communication system. Because it is not a must for word form to have a correlation to the meanings that they imply, this makes it much convenient for words to refer to abstract concepts. It would be difficult to restrict the form of signals to just the correlation between a symbol and an abstract meaning (Kuthy, 2001).
3.1.2 Linguistic structure – Discreteness
Thirdly, and contrastingly, the second set of structuring rules which combine groups of morphemes into words and sentences are semantically motivated. This is to say each language has a particular logic for combining such morphemes, so that its users are able to immediately figure out what each word, phrase or sentence specifically means, based on how the various morphemes are ordered. Accordingly, the fact that although language occurs as streams of continuous sounds or gestures, it can be analysed as individual units of meaning that are structured in systematic language-specific ways, helps to explain the core language feature of discreteness.
3.1.3 Linguistic semiotics – Displacement and Productivity
Fourthly, the co-working of these two sets of structuring rules helps language users communicate ideas about things which are not physically or temporally present, and indeed anything thing that they can clearly conceptualise. This is because at the fundamental level of sign, to use Fitch’s componential view, there is no inherent relationship between the signals of a language and the ideas they can represent, there is no restriction on the things that language users can communicate about – which helps explain the core language feature of displacement. Similarly, because morphemes can be further combined in various ways to form more complex meanings, and the number of possible combinations are practically infinite, languages can continue to expand the number of ideas within their communicative scope – which helps explains the core language feature of productivity.
3.3 Environment: Cultural Transmission
Fifthly, as a system of human communication, language is also a cultural tool. Accordingly, different human communities, living in different geographical areas, develop different cultures, of which their respective languages are part of. Further, according to Maynard Smith and Szathmary (1997), language is unique in being both part of human culture and a system for transmitting cultural information as well. Consequently, this helps explains the core language feature of cultural transmission, since humans need to acquire their native language from the communities they were born in – rare cases of children (e.g. the case of Genie) without such community support do not ever fully acquire language subsequently.
Children can learn from parents, human beings transmit their language down from one generation to another. A child brought up in isolation does not acquire language but birds reared in isolation still sing songs, although in very impoverished forms. (Chan, 2013)
2. Systematic Components
2.1 Language as a communication system
Any system of communication can be broken down into potentially three components: signs, structure, and semiotics. At the basic level, all systems comprise a collection of codes or signs. Optionally, some systems may prescribe methods or rules for ordering these signs in a structured manner. However, all systems include a conventionally established manner of interpreting its collection of codes, in other words a semiotics for these signs that has been agreed upon by its community of users.
With regards to language, formal linguistics has long divided this communication system into three such components. At the basic level, spoken and signed languages make use of language-specific stores of speech sounds (audio-vocal signs) or manual gestures (visuo-manual signs), which falls within the study of phonetics and phonology. All languages employ a system of rules for combining and ordering these signs into structured words and sentences, which falls within the study of morphology and syntax. Finally, within a community of language users there are conventionally established ways of interpreting these patterns of words and sentences, which falls within the study of semantics and pragmatics.
|
Figure 1: A componential view of language as a communication system |
|
| Communicative | Linguistic |
| Collection of signs | phonetics & phonology |
| Rules for structuring | morphology & syntax |
| Conventions of interpretation | semantics & pragmatics |
2.2 Language as a human cognitive faculty
In 2007, the evolutionary biologist William Tecumseh Fitch published an article on the evolution of language, in which he corresponded these three main components of language to three broad cognitive capacities underlying the human language faculty. In so doing, Fitch offered a view on the nature of language as a uniquely human communication system owing to its cognitive complexity across each of its three components.
2.2.1 Signal imitation
Natural languages have far larger stores of signs – traditionally, the number of consonant and vowel patterns in spoken languages – compared to other animal communication systems.
Accordingly, Fitch argues that humans must possess the cognitive capacity for complex, structured imitation – the starting point for a community for language users to build up a large lexicon of mutually agreed upon signs.
2.2.2 Structure generation and mapping
Natural languages have various systems of rules of great complexity and computational power which govern the combinatorial and hierarchical patternings of these signs.
Accordingly, Fitch argues that humans must possess the cognitive capacity to process such computationally sophisticated rules, particularly those responsible for the key property of recursion in syntax.
2.2.3 Semiotic drive
Natural languages have highly complex semiotic systems, in which the meanings of linguistic signs are dependent on and affected by their contexts of use. Crucially, language contrasts with other animal communication systems, in being a dedicated and intentional system for expressing and communicating ideas. For example, vervet monkeys instinctively produce alarm calls the moment they perceive nearby predators, however language users do not find themselves compelled to speak or sign due to similar external influences. Thus, when humans use language, they do so with full intention.
Accordingly, Fitch argues that humans must possess a cognitive appreciation of a “theory of mind”, unlike other animals. This means that language users understand that any conversation involves two or more distinct minds, and thus they use language as an intentional and specific means for creating or altering thoughts in the minds of their fellow language users, rather than under the influence of external stimuli.
|
Figure 2: Fitch’s tri-componential view of language |
||
| Communicative | Linguistic | Cognitive |
| Lexicon of Signifiers | phonetics & phonology | Signal imitation |
| Patterning/ Ordering rules | morphology & syntax | Structure generation and mapping |
| Signified meanings | semantics & pragmatics | Semiotic Drive |
2.3 Evaluation of Fitch’s components
Fitch’s analysis of the human language faculty helps us understand the various uniquely human cognitive capacities underlying language. However, in order to explain how these components interact with each other to form language-in-practice, as an empirical phenomenon, we feel that a dual approach combining both featural and componential analyses is necessary, which will be the topic of the following section.
